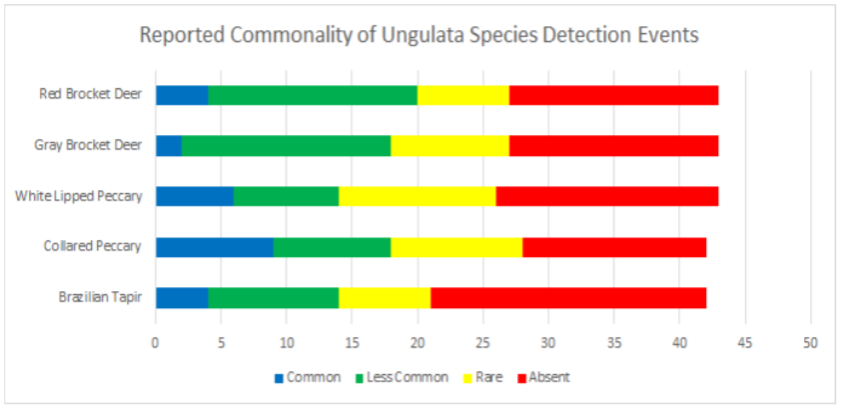
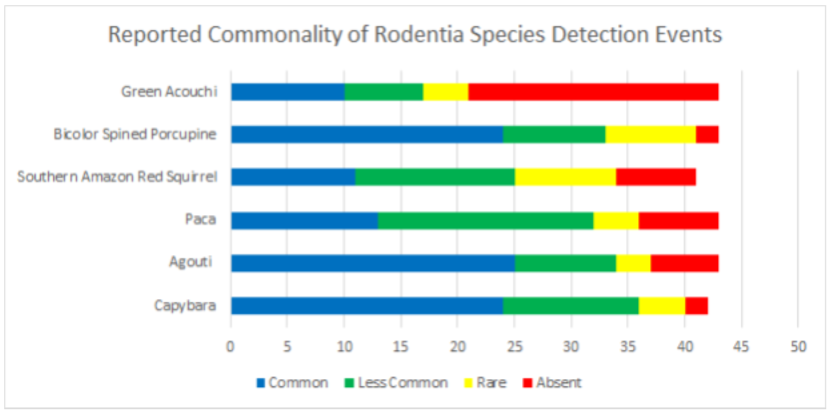
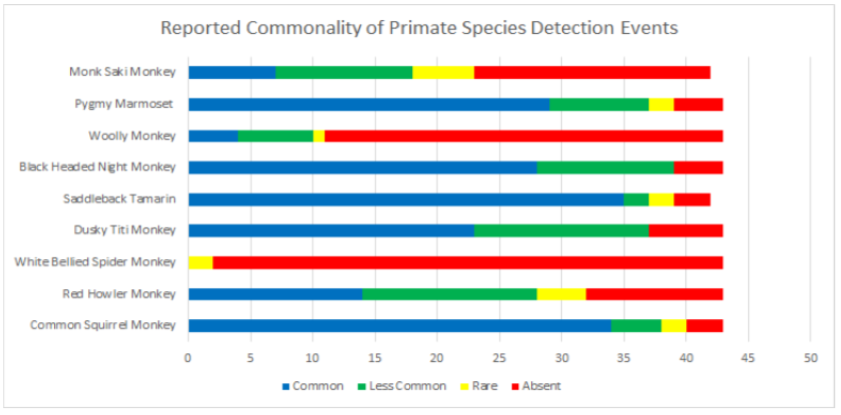
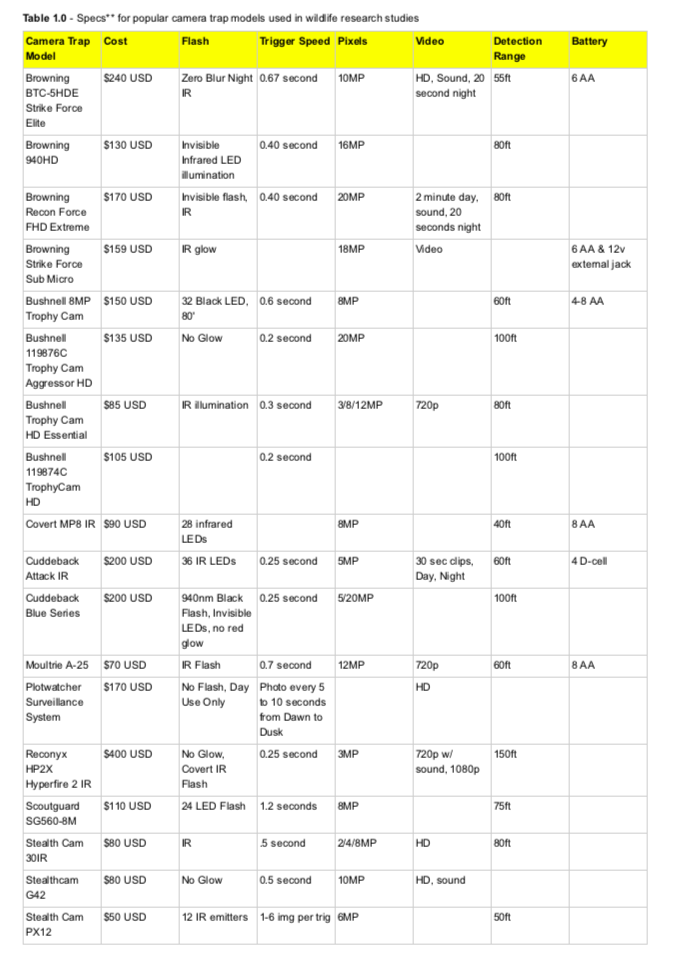
1.1 Abstract
This paper is a review and synthesis of medium (2-15 kg) to large (>15 kg) bodied Neotropical terrestrial mammals and their abilities to act as bioindicators of ecological health. As human behavior interferes with Neotropical forests for development, farm and cattle expansion, logging, and unsustainable hunting and harvesting, habitats may be clearcut and fragmented while animal populations may be extirpated, losing the important ecological services these species provided their ecosystem. By identifying these ecological services provided and monitoring and protecting these species’ populations, we can ensure healthy ecosystems into the future. Apex predators like jaguars and pumas help create a balance in prey species populations which trickles down to balancing plant biodiversity while also provisioning tertiary consumers. Tapirs and agoutis both act as crucial seed dispersers for large seeded Neotropical trees, aiding in colonizing new habitats and diversifying existing ones. Peccaries act as seed predators, balancing flora diversity while also protecting plants from insect predation and parasitism while also creating habitat for gap-dependent species and amphibians by wallowing and rooting behaviors. As these species vanish from an ecosystem, cascading effects can be seen by the loss of other species, both flora and fauna, dependent on their ecological services or complete shifts in the collective biodiversity of flora and fauna populations.
2.1 Introduction
Protecting large regions of neotropical rainforest has been considered a best practice for conserving biodiversity (Bruner et al., 2001). Nonetheless, protected rainforest areas may still suffer from the threat of human encroachment, unsustainable harvesting of flora and fauna, as well as logging (Laurance et al., 2012). In some cases, areas under protection have been so negatively impacted by human activities that a loss of ecological services has occurred (Jorge et al., 2013). While it is challenging to map ecological processes like community dynamics and nutrient cycling over large areas, an easier alternative exists through documenting and mapping specific species that play important ecological roles (Jorge et al., 2013) that include improving soil quality, dispersing of seeds, managing wildlife populations, altering herbivory, and mitigating wildfires and the spread of zoonotic diseases (Estes et al., 2011). With medium (2-15 kg) to large (> 15 kg) bodied terrestrial mammals disproportionately providing these ecological services (Novack et al., 2005; Estes et al., 2011; Campos-Arceiz et al., 2012), this paper will analyze the ways in which these species act as bioindicators for ecological health in neotropical forests and can aid in focusing prospective camera trap studies on species of significance.
3.1 Mammalian Apex Predators Influence (Panthera onca and Puma concolor)
Throughout much of the neotropics, two apex predator species exist - jaguars (Panthera onca) and pumas (Puma concolor). While avoiding each other temporally, these apex predators often overlap spatially in habitat use (Harmsen et al., 2009), and when doing so, their sympatric relationship triggers an evolutionary response in the puma population, making them smaller in physical size, likely to minimize competition with jaguars (Taber et al., 1997). Through comparative analysis of puma head and body length ratios across the species’ entire geographic range, Iriarte et al. (1990) found that pumas become smaller in size when sympatric with jaguars, likely correlating with jaguars taking larger prey (> 15 kg) and pumas taking medium and small prey (1-15 kg) (Maxit 2001; Polisar et al., 2003; Scognamillo et al., 2003; Azevedo 2008). Furthermore, when predators are sympatric with one another, they form a mammalian predator guild where they act as ecosystem architects (Terborgh et al., 2002), sustaining balance and structure within their environment’s food webs (Estes et al., 2011). As opportunistic generalists, jaguars and pumas feed upon large ungulates and various medium to small prey (Novack et al., 2005; Ruth & Murphy, 2010), keeping herbivore populations in check and preventing over consumption of flora (Terborgh, 1988; Ripple et al., 2016). Jaguars and pumas can further protect vegetation from herbivores by creating a landscape of fear, shifting behavioral patterns and habitat use, further reducing the consumption of plants in focused areas (Ford et al., 2014; Donadio & Buskirk, 2016). Through their influence, jaguars and pumas create a trophic cascade by controlling herbivorous prey populations, altering herbivory on the local flora, and creating a more diverse biomass of plant life which then can spawn more diversity in herbivorous and frugivorous fauna as more food niches are able to grow into maturity (Schmitz et al., 2000; Estes et al., 2011; Ripple et al., 2014).
Apex predators like jaguars and pumas also provide ecological services to nutrient cycling by influencing the spatial organization of herbivores (Bai et al., 2012; Schmitz et al., 2010). As herbivores influence nutrient pools in the soil by changing the configuration of plant populations through consumption, plant litter quantities decrease and alter the distribution of carbon and nutrients in plant tissues (Wardle et al., 2002; Bai et al., 2003). The landscape of fear apex predators instill upon these herbivores can further influence the way the prey species transport nutrients across habitats, fertilizing soils through feces and urine (Murray et al., 2013).
3.2 Mammalian Prey Species Influence
3.2.1 Tapirs (Tapirus)
As tropical rainforests house vast amounts of biodiversity, act as sources for carbon storage, and regulate climate, mitigating forest loss and degradation is vital (Lewis et al., 2015). While conservationists fight to protect large areas of neotropical forest, they are aided by the abilities of lowland tapirs (Tapirus terrestris) and Baird’s tapirs (Tapirus bairdii) to support forest regeneration and potentially offset disturbances that include habitat destruction and fragmentation, wildfires, and extreme climate events (Paolucci et al., 2018). As lowland tapirs are known to feed upon the fruits of about 300 plants (Barcelos et al., 2013) and cover home ranges up to 470 hectares (Tobler, 2008), this large terrestrial mammal plays a key role in seed dispersal, specifically of large seeds that smaller mammals are not as proficient at dispersing (Bueno et al., 2013; O’Farrill et al., 2013). This effectivity of dispersal is partly attributed to tapirs’ digestive systems only damaging between 1-31% (Hilbert et al., 2011; Paolucci et al., 2018) of ingested seeds as well as their preference for defecating in latrines. These accumulations of feces and seeds then act as hotspots for plant regeneration (Fragoso, 1997; Paolucci et al., 2018). Furthermore, tapirs aren’t just important at dispersing large seeds but also medium and small seeds as the majority of the seed load they pass through their gut has been observed to significantly favor smaller seeded species (Tobler et al., 2009).
Paolucci et al. (2018) also found that while tapirs have a great ability to spread seeds in healthy forest areas, they have an even greater ability to spread seeds in degraded forest areas as they often spread a mix of both pioneer and climax species as well as a mix of trees, shrubs, and lianas to these areas with minimal competition for light access. This is partly aided by the long gut retention rates (2-15 days) tapirs have in combination with their large home ranges, transporting seeds far away from their parent plant (O’Farrill et al., 2013). As tapirs defecate in latrines far from the ingested seed source, tapirs provide a fecal barrier, protecting seeds from seed predators and parasites while also providing fertilizer, increasing rates of germination (Fragoso 1997; Fragoso et al., 2003; O’Farril et al., 2013). This increased germination is further aided by other species like dung beetles transporting small seeds further away from the tapir latrines (Quiroga-Castro & Roldán, 2001; Rios & Pacheco, 2006) and medium to large sized seeds being further dispersed by scatter-hoarding rodents like agoutis (Brewer & Rejmanek, 1999).
3.2.2 Agoutis (Dasyprocta)
Agoutis (Dasyprocta), the medium sized (2-15 kg) terrestrial rodent species act as one of the aforementioned scatter-hoarders (Forget, 1994). While agoutis often live in high population densities (Wright et al., 1994), this in combination with their scatter-hoarding tendencies gives the species an important role as seed dispersers (Smythe, 1989; Asquith et al., 1999). When agoutis forage across their 2-3 ha area home ranges for fallen fruits or amongst the latrines of tapirs for seeds, agouti scatter-hoarding behavior leads them to act as short distance seed dispersers (Smythe, 1989; Fragoso, 1997) as most seeds are initially cached <25 m from their foraged locations (Jansen & Forget, 2001). Through this same scatter-hoarding behavior, they also collectively act as long distance seed dispersers by stealing from other agouti’s caches and re-caching their finds further away (Jansen & Forget, 2001; Haugaasen et al., 2010; Jansen et al., 2012). This act of secondary dispersal, usually within 5 to 200 m of their foraged location, partially fits within the often used long distance seed dispersal threshold of >100 m (Russo & Augspurger, 2004; Jordano et al., 2007; Jansen et al., 2012). Through this combination of short and long distance seed dispersal, not only do agoutis play a role into the aggregated distribution of large seeded tree species in Neotropical forests (Lieberman & Lieberman, 1994; Peres & Baider, 1997; Silvius & Fragoso, 2003), but also into the colonization of new habitats (Jansen et al., 2012). The dispersal efforts of agoutis prove to be so effective that even some species of trees, such as the Brazilnut tree (Bertholletia excelsa), are almost exclusively dependent on agoutis for the dispersal of their seeds (Peres & Baider, 1997; Taylor, 2000).
3.2.3 Peccaries (Tayassuidae)
In contrast to tapirs and agoutis acting as seed dispersers, peccaries (Tayassuidae) primarily play an important role as a seed predator (Beck, 2006). This important role peccaries play is clearly indicated when they are extirpated from a Neotropical region. When peccaries were absent, Wyatt and Silman (2004) observed significant increases in uneaten seeds between 5000-6000% for palm species like Iriartea deltoidea and Astrocaryum murumuru. This observation coincided with a 70% increase of Astrocaryum seedlings, creating a uniformity in local plant life that did not exist in the presence of peccaries (Silman et al., 2003). Furthermore, as collared peccaries (Pecari tajacu) and white-lipped peccaries (Tayassu pecari) forage in herds (Eisenberg and Redford, 1999) and consume over 200 species of fruits, seeds, and seedlings (Roldán and Simonetti, 2001; Beck, 2005), they provide forests with ecological services, creating balance in floral diversity through herbivory (Beck, 2006).
Peccaries also influence palm seedling establishment, spatial distribution, and density via trampling (Roldán and Simonetti, 2001). Trees with small seeded (< 1 cm diameter) fruits often survive peccary consumption, being dropped while eaten or surviving gut fermentation (Beck, 2005), eventually ending up trampled into the soil, providing a form of protection from insect seed predation (Silvius, 1999). Additionally, peccaries further aid seeds from insect predation as captive and field studies have both shown a dietary preference for seeds infested with larvae, likely for the added protein (Fragoso, 1994; Silvius, 2002). This insect predation by peccaries then acts as a mitigator to insect population increases and minimizes seed predators such as bruchid beetles’ impact on flora (Beck, 2006).
In addition to peccaries ecological interactions with seeds, they further act as ecosystem architects through rooting and wallowing behaviors (Jones et al., 1997). Both of these behaviors disturb leaf litter and soil of which act as chemical and physical barriers to litter-gap dependent species and the establishment of small seeded plant species, ultimately increasing flora diversity as these plant species now have access to habitat previously inaccessible (Metcalfe, 1996; Metcalfe & Turner, 1998, Lambert et al., 2005). The disturbance of leaf litter has also been shown to expose various insects to predation from vertebrate insectivores, aiding in the control of arthropod populations (Michel et al., 2014). Additionally, peccary wallows are also known to become vital habitats for breeding amphibians (Zimmerman and Simberloff, 1996; Reider et al., 2013), so much so that some amphibian species have been observed to go locally extinct soon after peccaries have become extirpated from an area (Simberloff, 1992).
4.1 Discussion
Large carnivores like jaguars and pumas serve as keystone species by providing ecological services of population management through direct predation, but also indirectly as food provisioners to scavengers (Wilmers et al., 2003). For this reason alone, jaguars and pumas act as a significant bioindicator for ecological health (Mora, 2017). Their presence demands a plethora of resources, ranging from abundant prey species to the vegetation and insects their prey species rely upon, to be largely intact. Their abilities to operate as ecosystem architects, sustaining balance and structure in local food chains, particularly in maintaining sustainable herbivory and mitigating the spread of zoonotic disease are soon felt upon their absence in an ecosystem (Terborgh, 1988; Terborgh et al., 2002; Estes et al., 2011; Ripple et al., 2016; Aguirre, 2017).
Tapirs’ abilities for long gut retention of seeds and ranging for long distances greatly aids in seed dispersal and in the diversification of flora amongst Neotropical forests (O’Farrill et al., 2013; Paolucci et al., 2018). This is particularly noticeable amongst large seeded (4-10cm in diameter) species of trees that once relied upon on the megafauna on the Pleistocene to disperse their seeds (Jansen et al., 2012). With these megafauna now absent, the tapir has taken up the role of long distance disperser of large seeds but is also aided by a much smaller mammal, the agouti (Jansen et al., 2012). While tapir latrines offer defecated seeds both fertilizer and protection from invertebrate parasites and seed predators (Fragoso 1997; Fragoso et al., 2003; O’Farril et al., 2013), they also provide agoutis access to seeds who then scatter-hoard their findings (Brewer & Rejmanek, 1999). While often dispersing seeds short distances as individuals, agoutis stealing from each others’ caches and re-caching the seeds elsewhere provide a species wide collective effort, functioning as a form of long distance seed dispersal as well (Smythe, 1989; Fragoso, 1997; Jansen & Forget, 2001; Haugaasen et al., 2010; Jansen et al., 2012). The combined efforts of tapirs and agoutis is clearly profound as they spread seeds of various sizes, diversifying both healthy and degraded forests (Silvius & Fragoso, 2003; Jansen et al., 2012; Paolucci et al., 2018). These combined efforts also favor arguments for reintroductions into areas where tapirs and agoutis have become extirpated as the loss of their ecological services has seen cascading effects in plant community structure, likely exacerbating global ecosystem services as early successional plant species over take dense wooded species that act as carbon sinks in these terrestrial mammal species’ absence (Bello et al., 2015; Kenup et al., 2018). Furthermore, tapirs and agoutis both have abilities to positively impact local economies through their seed dispersal services by promoting the establishment of various fruit species but also by acting as sources to bushmeat when hunted sustainably (Forget & Millerton, 1991; Taylor, 2000; Naidoo & Ricketts, 2006).
The importance of peccaries functioning as ecosystem architects is particularly evident in their absence. When extirpated from a region, the quantity of uneaten seeds rises significantly and plant diversity becomes more uniform in this seed predators’ absence (Silman et al., 2003; Wyatt and Silman, 2004). Plant uniformity may also be exacerbated by the loss of herbivory and trampling services provided by the social peccary species (Beck, 2006). Furthermore, with the peccaries’ ability to trample small seeds, protecting them from invertebrate seed predators as well as actively consuming invertebrate seed predator larvae, the absence of peccaries can lead to a loss in flora biodiversity through invertebrate plant predator infestations (Fragoso, 1994; Silvius, 2002; Silvius, 1999; Beck, 2006). Peccary absence also acts as a significant loss to apex predators like jaguars and pumas who then have to adapt to a major lack in prey source as well as amphibians who use peccary wallows as breeding habitats, all of which may follow the peccaries in extirpation if unable to adjust (Zimmerman and Simberloff, 1996; Sowls, 1997; Aranda, 2002; Reider et al., 2013). This loss can also be felt by humans as peccaries are important to the socio-economic status of humans living in forests due to peccaries acting as a provisional resource to meat and income (Bodmer et al., 1993; Stearman, 1992).
As jaguars, pumas, tapirs, agoutis, and peccaries act as keystone species to their ecosystem, the loss of their ecological interactions creates a significant threat to the ecosystem’s ability to function and remain structurally sound (Soule et al., 2003). Both the ecological and demographical extinction of these species can result in consequences that reach far and wide (Estes et al., 1989; Novaro et al., 2000) including the loss of biodiversity and the spread of zoonotic diseases (Jorge et al., 2013; Aguirre, 2017), making it imperative to effectively protect and monitor these populations. As these medium and large bodied terrestrial mammals are often cryptic in their nature and can live in challenging landscapes, trail cameras are optimal in gathering data to document species presence (Silver et al., 2004; Newey et al., 2015; Harmsen et al., 2017; Porfirio et al., 2018). While observing these species from firsthand accounts is rare, these devices offer a unique opportunity to document elusive species while preventing the habituation of animals to human presence (Borchers et al., 2014; Boyer-Ontl & Pruetz, 2014). Furthermore, collecting reliable population data on elusive species is needed to plan and implement proper conservation strategies in the practice (Nuñez-Pérez, 2011). Addiotionally, by engaging the local stakeholders who live amongst these neotropical species into conservation efforts, accurate knowledge can be learned, attitudes can change, and ecologically minded behaviors can develop through connections to wildlife species (Parsons et al., 2018) while collecting data can be made significantly less painstaking by recruiting and training local community members (Silver et al., 2004). By creating inclusive conservation programs, jaguars, pumas, tapirs, peccaries, agoutis, pacas, and more can survive largely in thanks to the people who live amongst these species, providing the ecosystem with their ecological services for generations to come.
5.1 Conclusions
Apex predators in the neotropics, such as jaguars and pumas, need abundant prey species to survive (Iriarte et al., 1990; Taber et al., 1997; Novack et al., 2005; Weckel et al., 2005; Jorge et al., 2013; Gutiérrez-González, 2017). Prey species like tapirs, agoutis, collared peccaries, and white lipped peccaries all aid in sustaining the large energy demands of these neotropical large felids as seasonal shifts occur (Novak et al., 2005; Weckel et al., 2006). Documenting the presence of these neotropical species assesses species richness and species density (Chetkiewicz et al., 2006; Boyd et al., 2007; Zanin et al., 2015) and further aids in the comprehensive assessment of ecological health by mapping species for their provided ecological services (Jorge et al., 2013).
As apex predators keep a balance in prey species populations, the prey species’ negative ecological impacts are kept to a minimum while their positive ecological impacts flourish (Terborgh et al., 2002; Estes et al., 2011). In combination, these ecological services include herbivore population management, provisional resourcing to tertiary consumers, mitigating the spread of zoonotic diseases, nutrient cycling, balancing herbivory, promoting seed dispersal, seed predation balancing flora, fighting seed parasitism and predation from insects, creating habitat for other species of flora and fauna to thrive, promoting flora diversity, aiding in fruit tree propagation including species effective at carbon sequestration, as well as providing local human populations access to bushmeat. Each species mentioned in this paper has a role to play and an ideal ecosystem would have a complete inventory of its native species in balance with one another to provide these collective ecological services. With each missing species, a service is lost, thus altering a forest’s dynamics for the other surviving species to adjust to or if incapable of adjusting, to vanish from the local ecosystem.
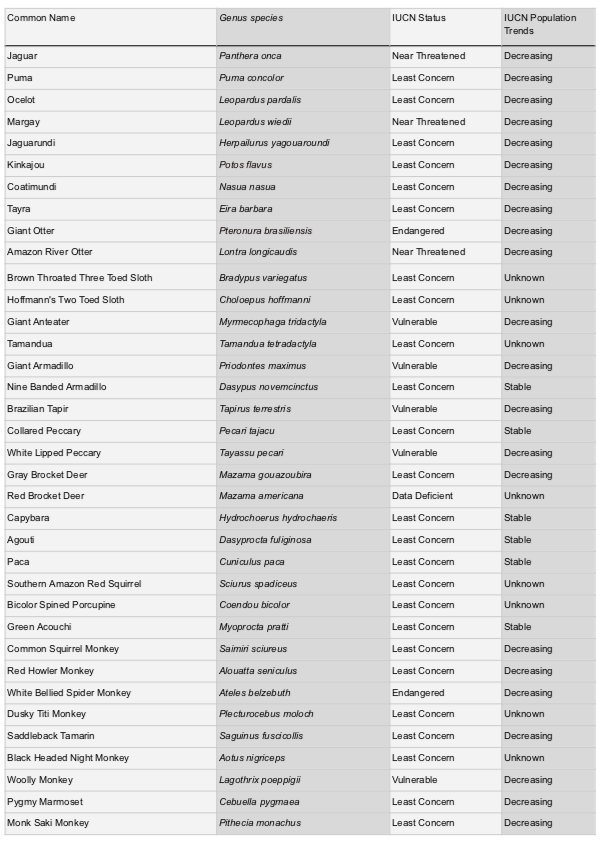
Investigations into additional terrestrial mammal species would be beneficial as well to further analyze the impact brocket deer (Mazama), pacas (Cuniculus), armadillos (Daspodidae), coatimundis (Nasua nasua), and others, giving a more complete understanding of Neotropical ecological services provided. Collectively, the species mentioned in this paper can effectively act as indicators of ecological health through their sustained presence in a region. Long term studies via trail camera data collection as well as sociological interviews would be beneficial to analyze species presence to create and maintain an understanding of the species’ ecological value to both the ecosystems and to the local human populations.
6.1 Works Cited
Aguirre, A. A. (2017). Changing patterns of emerging zoonotic disease in wildlife, domestic animals, and humans linked to biodiversity loss and globalization. ILAR Journal, 58(3): 315-318.
Aranada, M. (2002). Importancia de los pecaríes para la conservación del jaguar en México. In El Jaguar en el Nuevo Mileno. (Eds.) Medillin, R. A., Equihua, C., Chetkiewicz, C. L. B., Crawshaw Jr., P. G., Rabinowitz, A., Redford, K. H., et al., 101-106. Fonda de Cultura Económica, Ediciones Científicas Universitarias, Universidad Nacional Autónoma de México, and Wildlife Conservation Society, México.
Asquith, N. M., Terborgh, J., Arnold, A. E., & Mailen Riveros, C. (1999). The fruits the agouti ate: Hymenaea courbaril seed fate when its disperser is absent. Journal of Tropical Ecology, 15: 229-235.
Azevedo, F. C. C. (2008). Food habits and livestock depredation of sympatric jaguars and pumas in the Iguacu National Park area, south Brazil. Biotropica, 40: 494-500.
Bai, Y., Wu, J., Clark, C. M., Pan, Q., Zhang, L., Chen, S., … Han, X. (n.d.). Grazing alters ecosystem functioning and C:N:P stoichiometry of grasslands along a regional precipitation gradient. Journal of Applied Ecology, 49(6): 1204–1215. https://doi.org/10.1111/j.1365-2664.2012.02205.x
Barcelos, A. R., Bobroweic, P. E. D., Sanaiotti, T. M., & Gribel, R. (2013). Seed germination from lowland tapir (Tapirus terrestris) fecal samples collected during the dry season in the northern Brazilian Amazon. Integrative Zoology, 8: 63-73. https://doi.org/10.1111/1749-4877.12003
Beck, H. (2005). Seed predation and dispersal by peccaries throughout the Neotropics and its consequences: a review and synthesis. (Eds.) Forget, P. M., Lambert, J. E., Hulme, P. E., Vander Wall, S. B. Seed fate: predation, dispersal, and seedling establishment, 77-97. CABI Publishing, Wallingford, United Kingdom.
Beck, H. (2006). A review of peccary-palm interactions and their ecological ramifications across the Neotropics. Journal of Mammalogy, 87(3): 519-530.
Bello, C., Galetti, M., Pizo, M. A., Magnago, L. F. S., Rocha, M. F., Lima, R. A. F., et al. (2015). Defaunation affects carbon storage in tropical forests. Science Advances, 1: e1501105, https://doi.org/10.1126/sciadv.1501105.
Bodmer, R. E., Sowls, L. K., & Taber, A. B. (1993). Economic importance and human utilization of peccaries. In Status Survey and Conservation Action Plan: Pigs, Peccaries and Hippos: Status Survey and Conservation Action Plan. (Ed.) Oliver, W. L. V. 29-36
Borchers, D., Distiller, G., Foster, R., Harmsen, B., & Milazzo, L. (2014). Continuous-time spatially explicit capture-recapture models, with an application to a jaguar camera-trap survey. Methods in Ecology and Evolution, 5: 656-665.
Boyd, C., Brooks, T. M., Butchart, S. M., Edgar, G. J., Da Fonseca, G. B., Hawkins, F., & ... Van Dijk, P. (2008). Spatial scale and the conservation of threatened species. Conservation Letters, 1(1): 37-43.
Boyer-Ontl, K. M., & Pruetz, J. D. (2014). Giving the forest eyes: The benefits of using camera traps to study unhabituated chimpanzees (Pan troglodytes verus) in Southeastern Senegal. International Journal of Primatology, 35(5): 881-894.
Bruner, A. G., Gullison, R. E., Rice, R. E., & da Fonseca, G. A. B. (2001). Effectiveness of Parks in Protecting Tropical Biodiversity. Science, 291(5501): 125. Retrieved from https://search.ebscohost.com/login.aspx?direct=true&AuthType=cookie,ip&db=edsjsr&AN=edsjsr.3082189&site=eds-live&scope=site
Bueno, R. S., Guevara, R., Ribeiro, M. C. Culot, L., Bufalo, F. S., & Galetti, M. (2013). Functional redundancy and complementaries of seed dispersal by the last neotropical mega frugivores. PLoS One, 8, e56252. https://doi.org/10.1371/journal.pone.0056252
Brewer, S. W., and Rejmanek, M. (1999). Small rodents as significant dispersers of tree seeds in a Neotropical forest. Journal of Vegetation Science, 10: 165-174.
Campos-Arceiz, A., Traeholt, C., Jaffar, R., Santamaria, L., & Corlett, R. T. (2012). Asian Tapirs Are No Elephants When It Comes To Seed Dispersal. Biotropica, 44(2): 220. https://doi.org/10.1111/j.1744-7429.2011.00784.x
Chetkiewicz, C., St. Clair, C., & Boyce, M. (2006). Corridors for conservation: Integrating pattern and process. Annual Review of Ecology, Evolution, and Systematics, 37: 317-342.
Donadio, E., & Buskirk, S.W. (2016) Linking predation risk, ungulate antipredator responses, and patterns of vegetation in the high Andes. Journal of Mammalogy, 97: 966–977. doi:10.1093/jmammal/gyw020
Eisenberg, J. F., and Redford, K. H. (1999). Mammals of the Neotropics. The central neotropics: Ecuador, Peru, Bolvia, Brazil. University of Chicago Press, Chicago, Illinois, Vol. 3.
Elbroch, L. M., & Wittmer, H. U. (2012). Table scraps: inter-trophic food provisioning by pumas. Biology Letters, 8(5): 776–779. https://doi.org/10.1098/rsbl.2012.0423
Estes, J. A., Duggins, D. & Rathburn, G. (1989). The ecology of extinctions in kelp forest communities. Conservation Biology, 3: 252-264.
Estes, J. A., Terborgh, J., Brashares, J. S., Power, M. E., Berger, J., Bond, W. J. … David A. Wardle. (2011). Trophic Downgrading of Planet Earth. Science, 333: 301-306. https://doi.org/10.1126/science.1205106
Ford, A. T., Goheen, J. R., Otieno, T. O., Bidner, L., Isbell, L. A., Palmer, T. M., Ward, D., Woodroffe, R., Pringle, R. M. (2014). Large carnivores make savanna tree communities less thorny. Science 346: 346–349. doi:10.1126/science.1252753
Forget, P. M., and Millerton, T. (1991). Evidence for secondary seed dispersal by rodents in Panama. Oecologia, 87: 596-599.
Forget, P. M. (1994). Recruitment patterns of Vouacapoua americana (Caesalpiniaceae), a rodent-dispersed tree species in French Guiana. Biotropica, 26: 408-419.
Fragoso, J. M. V. (1994). Large mammals and the community dynamics of an Amazonian rain forest. Ph.D. thesis: University of Florida, Gainesville, Florida, United States.
Fragoso, J. M. V. (1997). Tapir-generated seed shadows: Scale-dependent patchiness in the Amazon rain forest. Journal of Ecology, 85: 519-529. https://doi.org/10.2307/2960574
Fragoso, J. M. V., Silvius, K. M., & Correa, J. A. (2003). Long-distance seed dispersal by tapirs increases seed survival and aggregates tropical trees. Ecology, 84: 1998-2006.
Gutiérrez-González, C. E., & López-González, C. A. (2017). Jaguar interactions with pumas and prey at the northern edge of jaguars’ range. PeerJ, 5, e2886.
Hairston, N. G., Smith, F. E., & Slobodkin, L. B. (1960). Community structure, population control, and competition. The American Naturalist, 94(879): 421-425.
Harmsen, B. J., Foster, R. J., Silver, S. C., Ostro, L. E. T., & Doncaster, C. P. (2009). Spatial and temporal interactions of sympatric jaguars (Panthera onca) and pumas (Puma concolor) in a Neotropical forest. Journal of Mammalogy, 903(3): 612-620.
Harmsen, B. J., Foster, R. J., Sanchez, E., Gutierrez-Gonzalez, C. E., Silver, S. C., Ostro, L. E. T., Kelly, M. J., Kay, E., & Quigley, H. (2017). Long term monitoring of jaguars in the Cockscomb Basin Wildlife Sanctuary, Belize; Implication for camera trap studies of carnivores. PLOS One, 12(6): 1-19.
Haugaasen, J. M. T., Haugaasen, T. Peres, C. A., Gribel, R., & Wegge, P. (2010). Seed dispersal of the Brazil nut tree (Bertholletia excelsa) by scatter-hoarding rodents in a central Amazonian forest. Journal of Tropical Ecology, 26: 251-262.
Hilbert, F., Sabatier, D., Andrivot, J., Scotti-Saintagne, C., Gonzalez, S., Prevost, M. F., Grenand, P., Chave, J., Caron, H., & Richard-Hansen, C. (2011). Botany, genetics and ethnobotany: a crossed investigation on the elusive tapir’s diet in French Guiana. PLoS ONE, 6(10): e25850.
Iriarte, J. A., Franklin, W. L., Johnson, W. E., & Redford, K. H. (1990). Biogeographic variation of food habits and body size of the America puma. Oecologia, 85 (2): 185–190.
Jansen, P. A., and Forget, P. M. (2001). Scatterhoarding and tree regeneration. (Eds.) Bongers, F., Charles-Dominique, P., Forget, P. M., & Thery, M. Nouragues: Dynamics and Plant-Animal Interactions in a Neotropical Rainforest, 275-288. Kluwer Academic, Dordrecht, The Netherlands, 275-288.
Jansen, P. A., Hirsch, B. T., Emsens, W-J., Zamora-Gutierrez, V., Wikelski, M., & Kays, R. (2012). Thieving rodents as substitute dispersers of megafaunal seeds. PNAS, 109(31): 12610-12615.
Jones, C. G., Lawton, J. H., & Shachak, M. (1997). Positive and negative effects of organisms as physical ecosystem engineers. Ecology, 78: 1946-1957.
Jordano, P., Garcia, C., Godoy, J. A., Garcia-Castano, J. L. (2007). Differential contribution of frugivores to complex seed dispersal patterns. PNAS, 104: 3278-3282.
Jorge, M. L. S. P., Galetti, M., Ribeiro, M. C., & Ferraz, K. M. P. M. B. (2013). Mammal defaunation as surrogate of trophic cascades in a biodiversity hotspot. Biological Conservation, 163: 49–57. https://doi-org.proxy.lib.miamioh.edu/10.1016/j.biocon.2013.04.018
Kenup, C. F., Sepulvida, R., Kreischer, C., & Fernandez, F. A. S. (2018). Walking on their own legs: unassisted population growth of the agouti Dasyprocta leporina, reintroduced to restore seed dispersal in an Atlantic Forest reserve. Oryx, 52(3): 571-578.
Laurance, W. F., Carolina Useche, D., Rendeiro, J., Kalka, M., Bradshaw, C. J. A., Sloan, S. P., … Coates, R. (2012). Averting biodiversity collapse in tropical forest protected areas. Nature, 489(7415): 290–294. https://doi.org/10.1038/nature11318
Lieberman, M., and Lieberman, D. (1994). Patterns of density and dispersion of forest trees. In McDade, L., Bawa, K. S., Hespenheide, H. A., & Hartshorn, G. S. (Eds.). La Selva: ecology and natural history of a Neotropical rain forest, 106-119. University of Chicago Press, Chicago, Illinois.
Lewis, S. L., Edwards, D. P., & Galbraith, D. (2015). Increasing human dominance of neotropical forests. Science, 349: 827-832. https://doi.org/10.1126/science.aaa9932
Maxit, I. E. (2001). Prey use by sympatric jaguar and puma in the Venezuelan llanos. M.S. Thesis: University of Florida, Gainesville, Florida, United States.
Metcalfe, D. J. (1996). Germination of small-seeded tropical rain forest plants exposed to different spectral compositions. Canadian Journal of Botany, 74: 516-520.
Metcalfe, D. J. and Turner, I. M. (1998). Soil seed bank from lowland rain forest in Singapore: canopy-gap and litter-gap demanders. Journal of Tropical Ecology, 14: 103-108.
Michel, N. L., Sherry, T. W., & Carson, W. P. (2014). The omnivorous collared peccary negates an insectivore-generated trophic cascade in Costa Rican wet tropical forest understory. Journal of Tropical Ecology, 30: 1-11.
Mora, F. (2017). A structural equation modeling approach for formalizing and evaluating ecological integrity in terrestrial ecosystems. Ecological Informatics, 41: 74-90.
Murray, B. D., Webster, C. R., & Bump, J. K. (2013). Broadening the ecological context of ungulate—ecosystem interactions: the importance of space, seasonality, and nitrogen. Ecology, 94(6): 1317. Retrieved from https://search.ebscohost.com/login.aspx?direct=true&AuthType=cookie,ip&db=edsjsr&AN=edsjsr.23436152&site=eds-live&scope=site
Naidoo, R. and Ricketts, T. H. (2006). Mapping the economic costs and benefits of conservation. PLoS Biology, 4(11): e360. https://doi.org/10.1371/journal.pbio.0040360.
Newey, S., Davidson, P., Nazir, S., Fairhurst, G., Verdicchio, F., Irvine, R. J., & Van der Wal, R. (2015). Limitations of recreational camera traps for wildlife management and conservation research: A practitioner’s perspective. Ambio, 44(4): 624-635.
Novack, A., Main, M., Sunquist, M., & Labisky, R. (n.d.). Foraging ecology of jaguar (Panthera onca) and puma (Puma concolor) in hunted and non-hunted sites within the Maya Biosphere Reserve, Guatemala. Journal of Zoology, 267: 167–178. https://doi.org/10.1017/S0952836905007338
Novaro, A. J., Funes, M. C., & Walker, R. S. (2000).Ecological extinction of native prey of a carnivore assemblage in Argentine Patagonia. Biological Conservation, 92: 25-33.
Nuñez-Pérez, R. (2011). Estimating jaguar population density using camera-traps: a comparison with radio-telemetry estimates. Journal of Zoology, 285: 39-45.
O’Farrill, G., Galetti, M., & Campos-Arceiz, A. (2013). Frugivory and seed dispersal by tapirs: An insight on their ecological role. Integrative Zoology, 8: 4-17. https://doi.org/10.1111/j.1749-4877.2012.00316.x
Paolucci, L. N., Pereira, R. L., Rattis, L., Silvério, D. V., Marques, N. C. S., Macedo, M. N., & Brando, P. M. (2018). Lowland tapirs facilitate seed dispersal in degraded Amazonian forests. Biotropica, 51: 245-252.
Parsons, A. W., Goforth, C., Costello, R., & Kays, R. (2018). The value of citizen science for ecological monitoring of mammals. PeerJ, 1-21.
Peres, C. A. (2005). Why We Need Megareserves in Amazonia. Conservation Biology, 19(3): 728. Retrieved from https://search.ebscohost.com/login.aspx?direct=true&AuthType=cookie,ip&db=edsjsr&AN=edsjsr.3591061&site=eds-live&scope=site
Peres, C., and Baider, C. (1997). Seed dispersal, spatial distribution and population structure of Brazilnut trees (Bertholletia excelsa, Lecythidaceae) in southeastern Amazonia. Journal of Tropical Ecology, 13: 595-616.
Polisar, J., Maxit, I., Scognamillo, D., Farrel, L., Sunquist, M. E., & Eisenberg, J. F. (2003). Jaguars, pumas, their prey base, and cattle ranching: ecological interpretations of a management problem. Biological Conservation, 109: 297-310.
Porfirio, G., Foster, V. C., Sarmento, P., & Fonseca, C. (2018). Camera traps as a tool for Carnivore conservation in a mosaic of Protected Areas in the Pantanal wetlands, Brazil. Nature Conservation Research: Zapovednaâ Nauka, 3 (2): 57-67.
Quiroga-Castro, V. D., and Roldán, A. I. (2001). The fate of Attalea phalerata (Palmae) seeds dispersed to a tapir latrine. Biotropica, 33: 472-477.
Reider, K. E., Carson, W. P., & Donnelly, M. A. (2013). Effects of collared peccary (Pecari tajacu) exclusion on leaf litter amphibians and reptiles in a Neotropical wet forest, Costa Rica. Biological Conservation, 163: 90-98.
Rios, R. S., and Pacheco, L. F. (2006). The effect of dung and dispersal on post-dispersal seed predation of Attalea phalerata (Arecaceae) by bruchid beetles. Biotropica, 38: 778-781.
Ripple, W. J., Estes, J. A., Beschta, R. L., Wilmers, C. C., Ritchie, E. G., Hebblewhite, M., Berger, J., Elmhagen, B., Letnic, M., Nelson, M. P., Schmitz, O. J., Smith, D. W., Wallach, A. D., & Wirsing, A. J. (2014). Status and ecological effects of the world’s largest carnivores. Science, 343: 124-1484.
Ripple, W. J., Estes, J. A., Schmitz, O. J., Constant, V., Kaylor, M. J., Lenz, A., … Wolf, C. (2016). What is a Trophic Cascade? Trends in Ecology & Evolution, 31(11): 842–849. https://doi.org/10.1016/j.tree.2016.08.010
Roldán, A. I., and Simonetti, J. A. (2001). Plant-mammal interactions in tropical Bolivian forests with different hunting pressures. Conservation Biology, 15: 617-623.
Russo, S. E., and Augspurger, C. K. (2004). Aggregated seed dispersal by spider monkeys limits recruitment to clumped patterns in Virola calophylla. Ecology Letters, 7: 1058-1067.
Ruth, T. K. & Murphy, K. 2010 Cougar –prey relations. Cougar: ecology and conservation (eds M. Hornocker & S. Negri), pp. 138 –162.
Schmitz, O. J., Hambäck, P. A., & Beckerman, A. P.. (2000). Trophic Cascades in Terrestrial Systems: A Review of the Effects of Carnivore Removals on Plants. The American Naturalist, 155(2): 141. https://doi.org/10.1086/303311
Schmitz, O. J., Hawlena, D., & Trussell, G. C. (n.d.). Predator control of ecosystem nutrient dynamics. Ecological Letters, 13(10): 1199–1209. https://doi.org/10.1111/j.1461-0248.2010.01511.x
Scognamillo, D. G., Maxit, I. E., Sunquist, M., & Polisar, J. (2003) Coexistence of jaguar (Panthera onca) and puma (Puma concolor) in a mosaic landscape in the Venezuelan llano. Journal of Zoological Society of London, 259: 269-279.
Silman, M. R., Terborgh, J. W., and Kilte, R. A. (2003). Population regulation of a dominant rain forest tree by a major seed predator. Ecology, 84: 431-438.
Silver, S. C., Ostro, L. E. T., Marsh, L. K., Maffei, L., Noss, A. J., Kelly, M. J., Wallace, R. B., Gomez, H., & Ayala, G. (2004). The use of camera traps for estimating jaguar Panthera onca abundance and density using capture/recapture analysis. Oryx, 38(2): 148-154.
Silvius, K. M. (1999). Interactions among Attalea palms, bruchid beetles, and neotropical terrestrial fruit-eating mammals: implications for the evolution of frugivory. Ph.D. Thesis: University of Florida, Gainesville, Florida, United States.
Silvius, K. M. (2002). Spatio-temporal patterns of palm endocarp use by three Amazonian forest mammals: granivory or “grubivory”? Journal of Tropical Ecology, 18: 707-723.
Silvius, K. M., and Fragoso, J. M. (2003). Home range use in an Amazonian forest: Implications for the aggregated distribution of forest trees. Biotopica, 35(1): 74-83.
Simberloff, D. (1992). Ecological communities. In McGraw Hill Encyclopedia of Science and Technology, 7(5). McGraw-Hill, New York, New York.
Smythe, N. (1989). Seed survival in the palm Astrocaryum standleyanum: Evidence for dependence upon its seed dispersers. Biotropica, 21: 50-56.
Soule, M., Estes, J. A., Berger, J., & Martinez Del Rio, C. (2003). Ecological effectiveness: conservation goals for interacting species. Conservation Biology, 17: 1238-1250.
Sowls, L. K. (1997). Javelinas and Other Peccaries: Their Biology, Management, and Use. 2nd ed. Texas A&M university Press, College Station, Texas, United States.
Stearman, A. M. (1992). Neotropical hunters and their neighbors: Siriono, Chimane and Yuqui hunting on the Bolivian frontier. In Conservation of Neotropical Forest. (Eds.) Redford, K., and Padoch, C. 108-128. Columbia University Press, New York, New York, United States.
Taber, A. B, Novaro, A. J., Neris, N., & Colman, F. H. (1997). The Food Habits of Sympatric Jaguar and Puma in the Paraguayan Chaco. Biotropica, 29(2): 204-213. Retrieved from https://search.ebscohost.com/login.aspx?direct=true&AuthType=cookie,ip&db=edsjsr&AN=edsjsr.2389036&site=eds-live&scope=site
Taylor, D. (2000). The Agouti’s nutty friend. International Wildlife, 30(2), 32.
Terborgh, J. (1988) The big things that run the world — a sequel to E.O. Wilson. Conservation Biology, 2 (4), 402–403.
Terborgh, J., Lopez, L., Nuñez V., P., Rao, M., Shahabuddin, G., Orihuela, G. … Balbas, L. (2002). Ecological Meltdown in Predator-Free Forest Fragments. Science, 294(5548): 1923. Retrieved from https://search.ebscohost.com/login.aspx?direct=true&AuthType=cookie,ip&db=edsjsr&AN=edsjsr.3085421&site=eds-live&scope=site
Tobler, M. W. (2008). The ecology of the lowland tapir in Madre de Dios, Peru: Using new technologies to study large rainforest mammals. PhD Dissertation, Texas A&M University. http://hdl.handle.net/1969.1/ETD-TAMU-2850.
Tobler, M. W., Janovec, J. P., & Cornejo, F. (2011). Frugivory and seed dispersal by the lowland tapir Tapirus terrestris in the Peruvian Amazon. Biotropica, 42(2): 215-222.
Wardle, D. A., Bonner, K. I., & Barker, G. M. (2002). Linkages between Plant Litter Decomposition, Litter Quality, and Vegetation Responses to Herbivores. Functional Ecology, 16(5): 585. Retrieved from https://search.ebscohost.com/login.aspx?direct=true&AuthType=cookie,ip&db=edsjsr&AN=edsjsr.826741&site=eds-live&scope=site
Weckel, M., Giuliano, W., & Silver, S. (2006). Jaguar (Panthera onca) feeding ecology: distribution of predator and prey through time and space. Journal of Zoology, 270: 25-30.
Wilmers, C. C., Crabtree, R. L., Smith, D. W., Murphy, K. M. & Getz, W. M. 2003 Trophic facilitation by introduced top predators: grey wolf subsidies to scavengers in Yellowstone National Park. Journal of Animal Ecology, 72: 909 –916. (doi:10.1046/j.1365-2656.2003.00766.x)
Wright, S. J., Gompper, M. E., & DeLeon, B. (1994). Are large predators keystone species in Neotropical forests? The evidence from Barro Colorado Island. Oikos, 71: 279-294.
Wyatt, J. L., and Silman, M. R. (2004). Distance-dependence in two Amazonian palms: effects of spatial and temporal variation in seed predator communities. Oecologia, 140: 26-35.
Zanin, M., Sollmann, R., Torres, N., Furtado, M., & Jácomo, A. T. A., Silveira, L., & De Marco Júnior, P. (2015). Landscapes attributes and their consequences on jaguar Panthera onca and cattle depredation occurrence. European Journal of Wildlife Research, 61(4): 529-537.
Zimmerman, B., and Simberloff, D. (1996). An historical interpretation of habitat use by frogs in a central Amazonian forest. Journal of Biogeography, 23(1): 27-46.
Adam J. Dewey, Miami University - Global Field Program, M.A. Biology