1.0 Abstract
Conservation management decisions need to be based on research evidence but when scientific data gaps exist in regions, local ecological knowledge (LEK) surveys can initially aid in filling this information void. Through LEK interviews, data was collected on 36 mammal species in 46 rural communities in the northeastern Peruvian Amazon tracking presence-absence, commonality, and 10 year population trends in October and November of 2019. Through data analysis, a statistically significant difference exists in the number of species present when comparing communities near the city of Iquitos, Peru and the Maijuna-Kichwa Regional Conservation Area (MKRCA). Species with low relative frequencies included white-bellied spider monkeys, woolly monkeys, and pumas. Species with reported decreasing populations included jaguars, pumas, ocelots, gray brocket deer, red brocket deer, and white-bellied spider monkeys. With the combined relative frequency and population trend data, there is likely reason for concern with these species and possible measures to protect them may be needed to prevent local extirpation events.
2.0 Introduction
Often times investigating cryptic wildlife can be exceptionally rigorous and expensive to study (Turvey et al., 2015; Martiez-Marti et al., 2016). As objective evidence plays an important role in conservation management decisions, a lack of data can lead to delays in the conservation actions needed to protect threatened habitats and species (Groombridge et al., 2004). While most ecological field research is directly observed and collected by professional and student scientists, other options exist. Utilizing local ecological knowledge (LEK) from communities who regularly interact with the natural habitats around them can garner a wealth of ecological data, particularly when gaps in data are present (Burbidge et al., 1988; Newing, 2011).
As LEK has gained an increasingly favorable view as a source for conservation research in vertebrates (Jones et al., 2008; Turvey et al., 2014), LEK interviews and surveys have also provided an inexpensive alternative for gathering comparative data in large scale and challenging landscapes (Turvey et al., 2015; Martinez-Marti et al., 2016). Despite these favorable views, LEK data still does not carry the scientific validity that conservation research performed by professional scientists does within the academic community and LEK data must be accepted as having a degree of uncertainty (Caruso et al., 2017). Regardless of this uncertainty, LEK is not rendered useless as proper data collection and analytical procedures minimize potential uncertainties while providing novel understanding into the state of various wildlife species and the condition of their habitats (Meijaard et al., 2011).
LEK research projects have provided valued conservation insights in various mammal species across the planet including saola populations in southeast Asia (Turvey et al., 2015), felids, elephants, ungulates, and primates in Equatorial Guinea (Martinez-Marti et al., 2016), and jaguars in Nicaragua (Zeller et al., 2011). These few examples are amongst many other LEK interview research projects that have gathered large swaths of data that were previously absent while also complimenting other scientific projects like camera trap and faecal DNA studies. This tactic of collecting LEK data is particularly valuable in research situations covering massive landscapes like wildlife corridors, areas with little to no prior research, regions with challenging landscapes, projects with limited time frames, and projects where budgets are constrained (Zeller et al., 2011; Martinez-Marti et al., 2016). This LEK data can then be analyzed for simple presence-absence within habitats but also identify population declines, particularly significant declines or even localized extirpation events (Strayer, 1999).
Through LEK survey-interviews, this paper will investigate the difference in 36 mammal species’ (See Appendix 9.1) presence-absence, commonality, and ten year population trends amongst 46 communities participating in CONAPAC’s (Conservacion de la Natureleza Amazonica del Peru, A.C.) Adopt-A-School program. Additionally, communities near the city of Iquitos will be compared to those near the Maijuna-Kichwa Regional Conservation Area (MKRCA), a known hub for wildlife propagation and preservation. CONAPAC has operated as an NGO (non-governmental organization) in the Peruvian Amazon since 1990 when a group of teachers, forestry engineers, and employees of the travel-lodging company, Explorama, created the organization in response to logging and oil exploration threatening the primary forests east of Iquitos, Peru (CONAPAC, n.d.). The Maijuna-Kichwa Regional Conservation Area (391,039 ha) was formed in 2015 after a ten year long effort by the indeginous community to have their ancestral territories and the native biodiversity protected by the Peruvian government (Roncal et al., 2018). Iquitos, the largest city in the Peruvian Amazon is a local economic hub grounded in timber, fisheries, oil, and tourism, all of which have varying elements that conflict with conservation interests in the region (Swierk & Madigosky, 2014; Orta-Martinez et al., 2018).
Currently, there is minimal mammal species data for the communities outside of the MKRCA, particularly many in the Adopt-A-School program and gathering a baseline set of data can provide information to base future conservation management decisions upon as the distribution of mammals would be better understood (Pillay et al., 2011). Due to the MKRCA having the same mammal species tracked for this LEK survey documented scientifically except for the white-bellied spider monkey (Gilmore et al., 2010), I predict that there will be a statistically significant difference where more mammal species are present near the MKRCA compared to the heavily human populated city of Iquitos. Furthermore, as the connectivity of forest landscapes plays a significant role in wildlife dispersal and promoting genetic diversity (Young and Clarke, 2000), I also expect to see a flow of species meandering through the partnered communities' forests towards the city of Iquitos out of the MKRCA during the mapping component of the action plan.
3.0 Materials and Methods
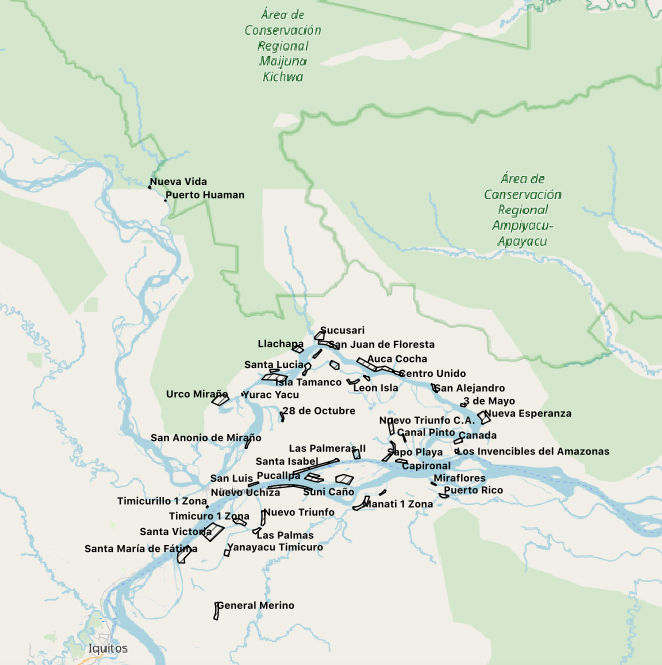
Community-based field work was conducted in the northeastern Peruvian Amazon (See Appendix 9.2; 9.3) during the annual evaluations for CONAPAC and the Detroit Zoological Society’s Adopt-A-School program during a two week period in October and November of 2019. Of the 55 partnered communities in the Adopt-A-School program, 46 were surveyed for mammal species presence, commonality, and sighting comparisons over the past 10 years (2009-2019). 24 communities reside along the Amazon river while another 23 reside along the Napo river, 2 along the Manati river, another 2 along the Yanayacu river, and 4 more along the Ucayal river (to be analyzed at a later date). The survey interviews were administered by staff from CONAPAC, the Detroit Zoological Society, as well as volunteers from the Peruvian Ministry of Environment and the Ministry of Education.
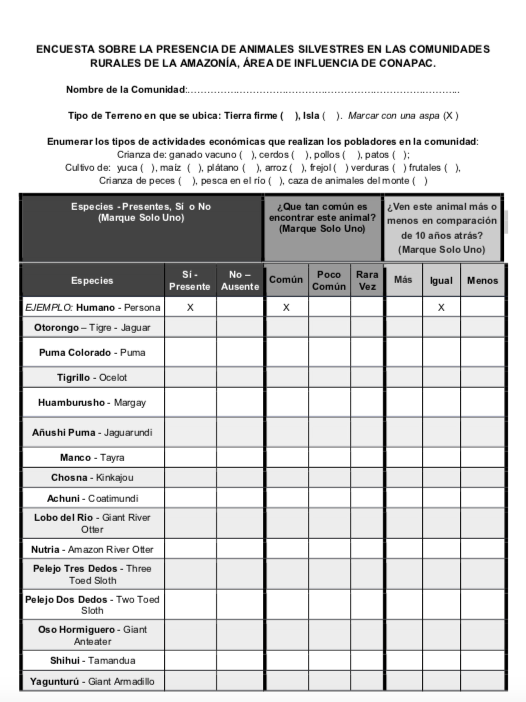
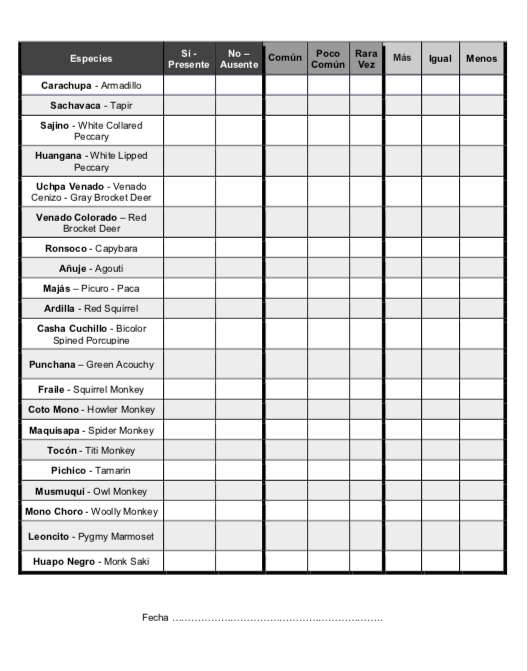
Data on the 36 mammal species was logged onto a survey (See Appendix 9.4) for easy data collection and to hold consistency across different survey administrators. This survey was accompanied by a statement providing the purpose of the study to prevent bias and garner participation from hunters and other individuals who would have an abundance of forest experience and knowledge to share. The survey included the species’ names in various overlapping regional dialects. Furthermore, to ensure accurate reporting, an animal ID card for the 36 mammal species surveyed was provided as a visual aid in identifying the different species present in the community forests. If an interviewer error of two checked boxes occurred in one category, no data would be collected for that species commonality and/or population trend. If this same error occurred in the presence-absence boxes, the species’ data would not be utilized by any means. When a community was not surveyed in 2019, 2018 data on presence/absence was utilized when available. Basic details on the community such as agricultural uses and mainland versus island community were also gathered.
This raw data was analyzed for various data sets. Presence-absence data was evaluated for relative frequency in the 46 surveyed communities. Data from 2019 was cross referenced to data from 2018 to analyze accuracy. Presence-absence data was then used within 16 communities - eight communities with the closest proximity to the city of Iquitos and eight communities with the closest proximity to the MKRCA to find statistical significance for these proximities and their role in the number of mammal species reported as present. This analysis utilized an unpaired t-test with an alpha value of 0.05. Furthermore, if mammal species were reported as present, they were then analyzed into reported commonalities that consisted of common, less common, rare, and absent categories and broken into different taxonomic groupings for ease (felidae, procyonidae, mustelidae, xenarthra, ungluta, rodentia, and primates). Population trend data reported by the 46 surveyed communities was also analyzed across all 36 mammal species. These data sets were then analyzed and graphed in Google Sheets and Excel.
4.0 Results
4.1 Statistical Analysis of Regional Influence on Mammal Species Presence
Two groupings of eight communities were broken into the categories of Iquitos and MKRCA communities and analyzed by an unpaired t-test. This tested confidence for the two community groupings’ proximity to their respective point of influence, the city of Iquitos and the MKRCA against the quantity of mammal species detected as present in those respective communities. The first grouping of eight communities were those in closest proximity to the city of Iquitos, Peru (General Merino, Las Palmas, Nuevo Triunfo C.A., Santa Maria de Fatima, Santa Victoria, Timicurillo, Timicuro I Zona, and Yanayacu Timicuro). The second grouping of eight communities were those in closest proximity to the MKRCA (Llachapa, Nueva Vida, Puerto Huaman, San Juan de Floresta, Santa Lucia, Sucusari, Urco Miraño, and Yurac Yacu Altura). Three of these communities are Maijuna communities bordering the MKRCA - Nueva Vida, Puerto Huaman, and Sucusari. An alpha of 0.05 was used to analyze the p-value of the t-tests. A p-value of 0.027 was obtained, rejecting the null hypothesis (no statistically significant difference) and thus showing the greater quantity of reported detection events in the MKRCA region than in the communities nearest to Iquitos to be statistically significant. This ultimately confirms my own hypothesis of proximity to the MKRCA playing a positive role in mammal species presence and proximity to the city of Iquitos having a negative role in mammal species presence.
4.2.1 Relative Frequency of Mammal Species
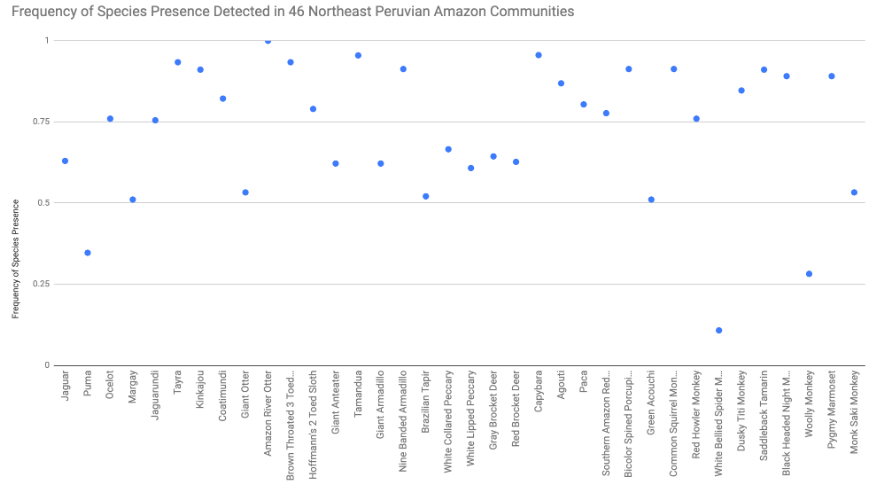
Relative frequency was utilized to show distribution for the 36 surveyed mammal species within the local environment of 46 communities (See Appendix 9.5). Three species were analyzed as having a frequency of < 0.500. These species include white-bellied spider monkeys (0.108), woolly monkeys (0.282), and pumas (0.347). Five species also had relative frequencies near 0.500 - green acouchis (0.511), margays (0.511), Brazilian tapirs (0.521), giant otters (.533), and monk saki monkeys (0.533). Ten species had relative frequencies of over 0.900 with one species reaching a relative frequency of 1.00 - kinkajou (0.911), saddleback tamarins (0.911), common squirrel monkeys (0.913), bicolor spined porcupines (0.913), nine-banded armadillos (0.913), tayras (0.934), brown-throated three toed sloths (0.934), tamanduas (0.955), capybaras (0.956), and Amazon river otters (1.00). The remaining 18 species had relative frequencies between 0.600 and 0.900. When relative frequency was further categorized into river systems (Amazon, Manati, Napo, and Yanayacu), two species were shown to only be reported as present in one river system. Pumas and white-bellied spider monkeys were only reported present along the Napo river communities (See Appendix 9.6; 9.7). Furthermore, 33 species showed relative frequencies for all four river systems and one species (wooly monkey) was present in three of the four river systems (Amazon, Manati, and Napo).
4.2.2 Reported Commonality of Species Detection Events
When species were reported present, their commonality was tracked into three main categories - common, less common, and rare (See Appendix 9.8). Absence was also utilized within graphs to aid the visualization of 100% of collected data. Of the 36 mammal species surveyed, 14 were reported as common in over 50% of community surveys (kinkajou, coatimundi, tayra, amazon river otter, brown-throated three toed sloth, tamandua, capybara, agouti, bicolor spined porcupine, common squirrel monkey, dusky titi monkey, saddleback tamarin, black-headed night monkey, and pygmy marmoset).
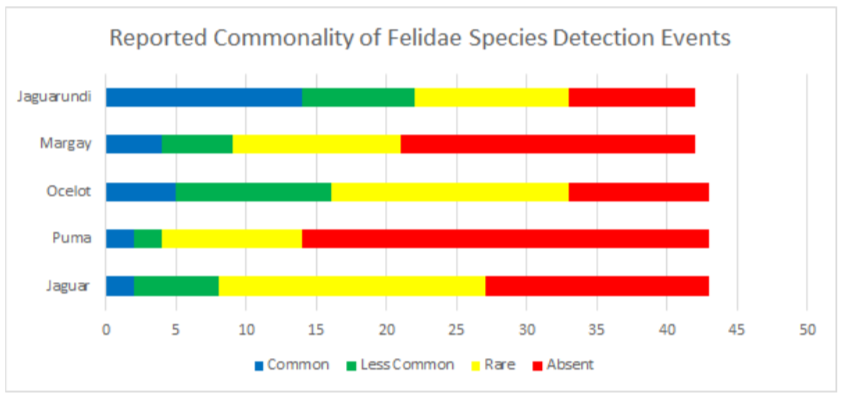
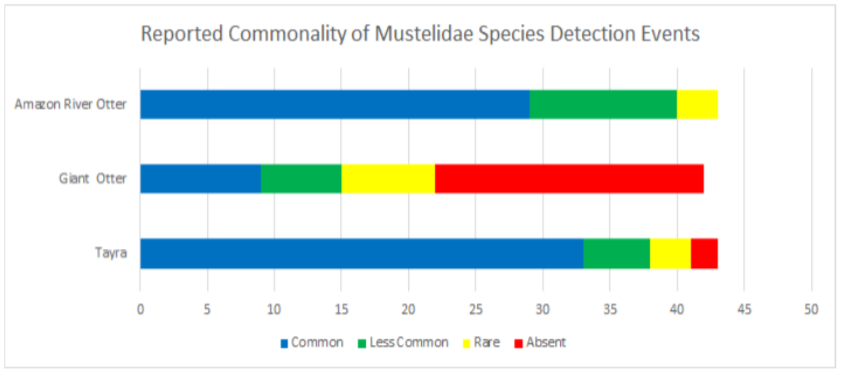
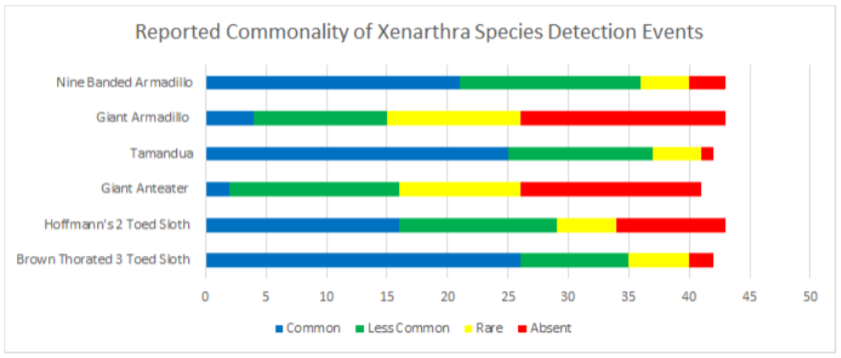
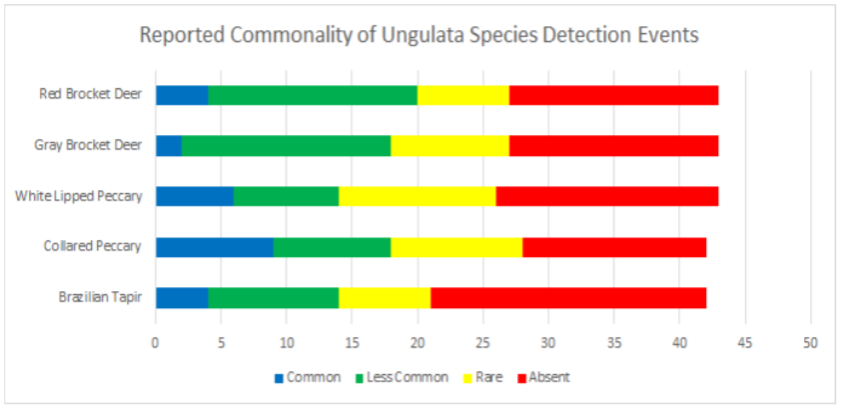
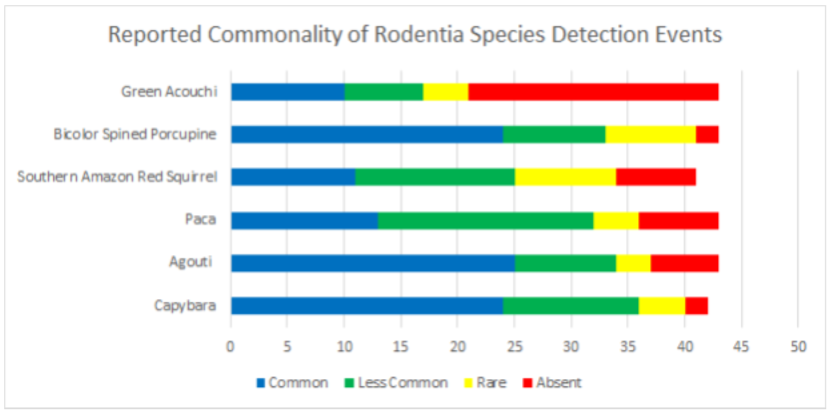
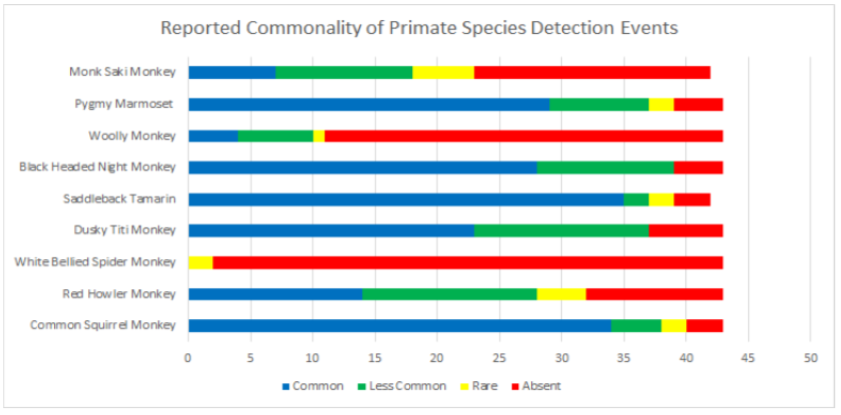
Data was also broken into different taxonomic classifications to see how species of similar evolutionary history may fair similarly or differently. When looking into the family felidae (See Appendix 9.9), jaguarundis were the most commonly seen species followed by ocelots, margays, jaguars, and lastly, pumas which were reported absent in 67.4% and rare in 23.2% of communities. Within the family procyonidae (See Appendix 9.10), both kinkajous and coatimundis were reported as commonly seen in over 50% of the communities. Furthermore, kinkajous were reported as as slightly more common than coatimundis. Of the mustelidae family (See Appendix 9.11) surveyed, tayras were the most common followed by the Amazon river otter. The giant otter was the least common of the mustelids with it being absent in 47.6% and rare in 16.6% of communities. Within the superorder xenarthra (See Appendix 9.12), brown-throated three toed sloths were reported as most common, respectively followed by tamanduas, nine-banded armadillos, Hoffmann’s two toed sloths, giant armadillos, and finally, giant anteaters. Giant armadillos were reported absent in 39.5% and rare in 25.5% of communities while giant anteaters were reported as being absent in 36.5% and rare in 24.3% of communities. Of the superorder ungulata (See Appendix 9.13), the collared peccary was reported as the most common followed by the white lipped peccary, tapir & red brocket deer, and finally the gray brocket deer. No single ungulate species was primarily reported as common, favoring reports of being less common, rare, or absent. Most notably, within surveyed communities, tapirs were reported absent in 50% of the time, white lipped peccaries were reported absent 39.5% of the time, red and gray brocket deer were reported absent 37.2% of the time each, and collared peccaries were reported absent 33.3% of the time. Within the order rodentia (See Appendix 9.14), agoutis, capybaras, and bicolored spined porcupines were the most common in this respective order. This was followed up by pacas and southern Amazon red squirrels favoring reports of being less common and the green acouchi favoring reports of absence in 51.1% of communities. For the order primate (See Appendix 9.15), saddleback tamarins were reported as most common followed by common squirrel monkeys, pygmy marmosets, black-headed night monkeys, and dusky titi monkeys. Red howler monkeys had equal reporting of common and less common detection events. The most notable data within primate commonality reporting pertains to white-bellied spider monkeys being absent in 95.3% of communities, woolly monkeys being absent in 74.4% of communities, and monk saki monkeys being absent in 45.2% of communities.
4.2.3 Mammal Species Population Trends Over the Past 10 Years as Reported by Community Members
When only analyzing communities that reported presence, population trends showed species as either increasing, stable, or decreasing (See Appendix 9.16). Of the species reported present, 25 were reported as increasing. These increasing populations included jaguarundis, tayras, kinkajous, coatimundis, giant otters, Amazon river otters, brown-throated three toed sloths, Hoffmann’s two toed sloths, tamanduas, nine-banded armadillos, collared peccaries, white-lipped peccaries, capybaras, agoutis, pacas, southern Amazon red squirrels, bicolor spined porcupines, green acouchis, common squirrel monkeys, red howler monkeys, dusky titi monkeys, saddleback tamarins, black-headed night monkeys, pygmy marmosets, and monk saki monkeys. Four present species were reported equally as increasing and stable - giant anteaters, giant armadillos, Brazilian tapirs, and woolly monkeys. Margays stood alone as the only present species most commonly reported as trending towards stable (without another categorical tie). The remaining six species had populations that were reported as decreasing - jaguars, pumas, ocelots, gray brocket deer, red brocket deer, and white-bellied spider monkeys. When factoring in absence, 15 species were most commonly reported with their population’s trend trending toward being absent - jaguars, pumas, margays, giant otters, giant anteaters, giant armadillos, Brazilian tapirs, collared peccaries, white-lipped peccaries, gray brocket deer, red brocket deer, green acouchis, white-bellied spider monkeys, woolly monkeys, and monk saki monkeys.
4.3 Agricultural Data
The presence of agricultural practices and the number of mammal species reported present in Adopt-A-School communities were correlated (See Appendix 9.17) to find if relationships were positive, negative, or if no relationship existed at all. Positive relationships between the number of mammal species present and the particular agricultural uses include - hunting, pigs, chickens, cattle, plantains, fish farms, wild caught fish, ducks, and corn in this respective order. It is important to note that no positive relationships surpassed a coefficient of 0.30 suggesting a weak positive relationship. Negative relationships included rice, beans, vegetables, fruits, and yuca. Similar to the positive relationships, no negative relationship surpassed a coefficient of -0.30 suggesting a weak negative relationship between the agricultural uses and presence of mammal species.
5.0 Discussion
The results of this LEK data collection on mammal species detection events provided extensive insights into the local population of mammals between Iquitos, Peru and the MKRCA. While it is difficult to validate without uncertainty, baseline data in Gilmore et al. (2010) provides a comparative set of information for what a healthy mammal ecosystem looks like in the region. Additionally, it is important to acknowledge that this study is a representation of detection events reported by community members - many hunters, farmers, and/or local authorities who carry valuable LEK but are also human and prone to lapses in memory. Due to these uncertainties, this data should be utilized as complementary data to other scientific methodologies such as camera trap and faecal DNA studies.
5.1 Regional Insights
The p-value analyzing mammal presence near Iquitos and the MKRCA confirms a statistically significant difference between the present populations. Much of the credit for the greater quantity of reported mammal species in the MKRCA region should go to the Maijuna communities and their hard work getting their ancestral lands and native biodiversity federally protected. This is particularly evident as the three Maijuna Adopt-A-School communities bordering the MKRCA were among the communities with the highest reported presence of mammal species - Nueva Vida with 97.2% of species and both Puerto Huaman and Sucusari with 94.4% of species surveyed. Furthermore, this mirrors the research in Gilmore et al. (2010). It should also be noted that during the creation of the MKRCA, the Maijuna retained their rights to hunt on ancestral land (Roncal et al., 2018). With the MKRCA being a sanctuary for wildlife, it would be valuable for future studies to analyze hunting methods in Maijuna communities in comparison with those outside of the immediate MKRCA region to better understand the varying hunting pressures being applied to different mammal species.
5.2 Species of Concern
Interview data around carnivores should be used with caution due to the high potential for bias (Caruso et al., 2016). These biases may be due to various reasons ranging from the charisma of a particular species, to current humans wildlife conflict issues (Turvey et al., 2015), to the cryptic behaviors of carnivores adding the likelihood of false-negatives where they are reported as absent when possibly present (Caruso et al., 2016; Martinez-Marti et al., 2016). Regardless of these potential biases, according to this study’s collected data, pumas (relative frequency of 0.347) appear to be absent everywhere except along Napo river communities. This could likely be due to a lack of prey base (Miotto et al., 2011), habitat degradation and human wildlife conflicts (Weber & Rabinowitz, 1996), or false negative reporting (Turvey et al., 2015) in the other river system regions. Giant otters (relative frequency of 0.533) were reported absent from nearly half of all communities. In the past, giant river otters faced intense hunting pressures (Gilmore et al., 2010), likely due to human wildlife conflicts where communities retaliated for fishing competition (Lima et al., 2014). Gilmore et al. (2010) shares further concerns of contamination from oil extraction also impacting giant otters. Giant otters populations may still be recovering from these different human impacts today. White-bellied spider monkeys and woolly monkeys were absent from most communities surveyed with respective relative frequencies of 0.108 and 0.282. According to Gilmore et al. (2010), white-bellied spider monkeys have been found to be absent in the MKRCA. Furthermore, white-bellied spider monkeys were also found absent in the nearby Ampiyacu-Apayacu Regional Conservation Area (AARCA) (Montenegro and Escobedo, 2004). As the absence of this species is usually attributed to hunting pressures too intense for the species to thrive (Aquino & Encarnación, 2004; Emmons & Feer, 1997), the data from this study likely confirms this hunting pressure is still too much for the large bodied primate species to radiate outward. This is probably a similar situation for woolly monkeys too as their relatively large bodies also make them a desirable source of bushmeat (Di Fiore, 2004).
Gilmore et al. (2010) suggests that implementing controls on the hunting of species with low reproductive rates (i.e. Brazilain tapir, woolly monkey, red howler) would aid medium and large bodied mammal population recovery as well as protect vital seed dispersers in the local environment (Stevenson et al., 2002; Gilmore et al., 2010; Bueno et al., 2013; O’Farrill et al., 2013). This would also be beneficial for the gray and red brocket deer in order to protect their complementary ecological roles as seed dispersers (Bodmer, 1991) and as their population trends appear to be decreasing. These hunting controls may further indirectly aid the carnivore populations with decreasing trends as well. As the prey base for jaguars and pumas are protected, the apex predators can inhabit more territory while providing various ecological services like mitigating disease vectors and diversifying flora through controlling herbivorous prey populations (Terborgh, 1988; Terborgh et al., 2002; Estes et al., 2011; Ripple et al., 2016; Aguirre, 2017).
5.3 Future Studies
While data from 2019 was cross referenced to data from 2018 to analyze accuracy, providing a consistent rating that stood at 88.33%, additional layers could be taken to insure accuracy and mitigate bias in future studies. One added measure suggested by Zeller et al. (2011) and Petracca et al. (2017) is to vet out false-positive reports by providing a brief test of sorts, where the interviewee would first identify pictures and tracks of species to show reliability in their accurate identification of the species being researched. In this paper’s study, the animal ID card operated in a similar manner and aided in clarifying species with similar appearances like the ocelot and margay and/or those long extirpated in localized regions where local species names became interchangeable like with the amazon river otter (nutria) and the giant river otter (lobo del rio) in some communities. It may also be beneficial to include multiple hunter interviews per location to provide added cross referencing. Additionally, continuing LEK surveys in the years to come would monitor changes in species’ distribution and frequency over time.
This study will include data from four additional communities along the Ucayali river near the Tamshiyacu-Tahuayo Regional Conservation Area in the future.
6.0 Conclusions
While 28 mammal species have relative frequencies of over 0.600 and 25 mammal species have reported population trends as increasing, other species are not faring so well. With eight mammal species having relative frequencies of 0.533 or less - (white-bellied spider monkeys [0.108], woolly monkeys [0.282], pumas [0.347], green acouchis [0.511], margays [0.511], Brazilian tapirs [0.521], giant otters [.533], and monk saki monkeys [0.533]), and six species with populations reported as decreasing - (jaguars, pumas, ocelots, gray brocket deer, red brocket deer, and white-bellied spider monkeys), there is a reason for concern as many of these species play a significant ecological role as ecosystem architects through seed dispersal, population management, disease mitigation, and more. Scientific research into these populations would be beneficial to see if the LEK data aligns. With this scientific data, proper conservation management decisions could be put forth to protect the species of concern while incorporating the people who live amongst them. In the meantime, a focussed emphasis on the specific species of concern within education programing would aid in creating awareness of these issues and further cultivate sustainable minded attitudes and behaviors (Feilen et al., 2017).
7.0 Acknowledgements
I would like to thank every participating community member in the Adopt-A-School program that shared their local ecological knowledge, making this research project possible. Fieldwork assistance was made possible by CONAPAC and supporting volunteers in efforts to collect this data; Claire Lannoye-Hall, Curator of Education at the Detroit Zoological Society for continuing to support me in both my academic and professional endeavors. To the Miami University professors and fellow students for continued support in the design of this research project.
8.0 Citations
Aguirre, A. A. (2017). Changing patterns of emerging zoonotic disease in wildlife, domestic animals, and humans linked to biodiversity loss and globalization. ILAR Journal, 58(3): 315-318.
Aquino, R., and Encarnación, F. (1994). Primates of Peru. Primate Report, 40: 1–127.
Bodmer, R. (1991). Strategies of Seed Dispersal and Seed Predation in Amazonian Ungulates. Biotropica, 23(3), 255-261. doi:10.2307/2388202
Bueno, R. S., Guevara, R., Ribeiro, M. C. Culot, L., Bufalo, F. S., & Galetti, M. (2013). Functional redundancy and complementaries of seed dispersal by the last neotropical mega frugivores. PLoS One, 8, e56252. https://doi.org/10.1371/journal.pone.0056252
Burbidge, A. A., Johnson, K. A., Fuller, P. J., & Southgate, R. J. (1988). Aboriginal knowledge of the mammals of the central deserts of Australia. Australian Wildlife Research, 5, 9-39.
Caruso, N., Luengos Vidal, E., Guerisoli, M., & Lucherini, M. (2017). Carnivore occurrence: Do interview-based surveys produce unreliable results? Oryx, 51(2): 240-245.
CONAPAC (n.d.). Who we are. CONAPAC Website. Retrieved from: https://www.conapac.org/who_we_are.html
Di Fiore, A. (2004). Primate conservation. Pp. 274–277 in McGraw-Hill Yearbook of Science and Technology. McGraw-Hill Companies, New York.
Emmons, L. H., and Feer, F. (1997). Neotropical Rainforest Mammals. The University of Chicago Press, Chicago.
Estes, J. A., Terborgh, J., Brashares, J. S., Power, M. E., Berger, J., Bond, W. J. … David A. Wardle. (2011). Trophic Downgrading of Planet Earth. Science, 333: 301-306. https://doi.org/10.1126/science.1205106
Feilen, K., Guillen, R., Vega, J. & Savage, A. (2017). Developing successful conservation education programs as a means to engage local communities in protecting cotton-top tamarins ( Saguinus oedipus ) in Colombia. Journal for Nature Conservation, 41. doi: 10.1016/j.jnc.2017.10.003.
Gilmore, M. P., Vriesendorp, C., Alverson, W. S., del Campo, A., von May, R., Lopez Wong, C., Rios Ochoa, S. (2010). Peru: Maijuna - Rapid Inventories: Biological and Social. The Field Museum, Chicago, IL, USA. pg. 312-320.
Groombridge, J. J., Massey, J. G., Bruch, J. C., Malcom, T., Brosis, C N., Okada, M. M., Sparklin, B., Fretz, J. S., VanderWerf, E. (2004). An attempt to recover the po’o-uli by translocation and an appraisal of recovery strategy for bird species of extreme rarity. Biological Conservation, 118, 365-375.
Jones, J. P. G., Andriamarovololona, M. M., Hockley, N., Gibbons, J. M., & Milner-Gulland, E. J. (2008). Testing the use of interviews as a tool for monitoring trends in the harvesting of wild species. Journal of Applied Ecology, 45, 1205-1212.
Lima, D. d. S., Marmontel, M., & Bernard, E. (2014). Conflicts between humans and giant otters (Pteronura brasiliensis) in Amanã Reserve, Brazilian Amazonia. Ambiente & Sociedade, 17(2), 127-142. https://dx.doi.org/10.1590/S1414-753X2014000200009
Martinez-Mart, C., Jimenez-Franco, M. V., Royle, J. A., Palazon, J. A. & Calvo, J. F. (2016). Integrating occurrence and detectability patterns based on interview data: a case study for threatened mammals in Equatorial Guinea. Scientific Reports, 6:3383. DOI: 10.1038/srep33838.
Meijaard, E., Buchori, D., Hadiprakarsa, Y., Utami-Atmoko, S. S., Nureahyo, A., Tiju, A. et al. (2011). Quantifying killing of orangutans and human-orangutan conflict in Kalimatan, Indonesia. PLoS One, 6, e27491.
Miotto, R. A., Cervini, M., Figueiredo, M. G., Begotti, R. A. & Galetti Jr., P. M. (2011). Genetic diversity and population structure of pumas (Puma concolor) in southeastern Brazil: implications for conservation in a human-dominated landscape. Conservation Genetics, 12: 1447-1455.
Montenegro, O., and Escobedo, M. (2004). Mamíferos/Mammals. Pp. 80–88 and 164–171 in Vriesendorp, C., Pitman, N., Foster, R., Mesones, I. & Rios, M., eds. Perú: Ampiyacu, Apayacu, Yaguas, Medio Putumayo. Rapid Biological Inventories Report 12. The Field Museum, Chicago.
Newing, H. (2011). Conducting Research in Conservation: A Social Science Perspective. Routledge, Abingdon, UK.
O’Farrill, G., Galetti, M., & Campos-Arceiz, A. (2013). Frugivory and seed dispersal by tapirs: An insight on their ecological role. Integrative Zoology, 8: 4-17. https://doi.org/10.1111/j.1749-4877.2012.00316.x
Orta-Martínez, M., L. Pellegrini, & M. Arsel. (2018). “The squeaky wheel gets the grease”? The conflict imperative and the slow fight against environmental injustice in northern Peruvian Amazon. Ecology and Society, 23(3):7. https://doi.org/10.5751/ES-10098-230307
Petracca, L. S., Frair, J. L., Cohen, J. B., Calderon, A. P., Carazo-Salazar, J., Castaneda, F., … Quigley, H. (2017). Robust inference on large-scale species habitat use with interview data: The status of jaguars outside protected areas in Central America. JOURNAL OF APPLIED ECOLOGY, 55(2), 723–734. https://doi-org.proxy.lib.miamioh.edu/10.1111/1365-2664.12972
Pillay, R., Johnsingh, A. J. T., Raghunath, R. & Madhusan, M. D. (2011). Patterns of spatiotemporal change in large mammal distribution and abundance in the southern Western Ghats, India. Biological Conservation, 144: 1567-1576.
Ripple, W. J., Estes, J. A., Schmitz, O. J., Constant, V., Kaylor, M. J., Lenz, A., … Wolf, C. (2016). What is a Trophic Cascade? Trends in Ecology & Evolution, 31(11): 842–849. https://doi.org/10.1016/j.tree.2016.08.010
Roncal, C. M., Bowler, M. & Gilmore, M. (2018). The ethnoprimatology of the Maijuna of the Peruvian Amazon and implications for primate conservation. Journal of Ethnobiology and Ethnomedicine, 14: 19. Doi: 10.1186/s13002-018-0207-x
Stevenson, P.R., Castellanos, M.C. & Pizarro, J.C. et al.(2002). Effects of Seed Dispersal by Three Ateline Monkey Species on Seed Germination at Tinigua National Park, Colombia. International Journal of Primatology, 23: 1187. https://doi.org/10.1023/A:1021118618936
Strayer, D. L. (1999). Statistical power of presence-absence data to detect population declines. Conservation Biology, 13(5): 1034-1038.
Swierk, L., & Madigosky, S. R. (2014). Environmental Perceptions and Resource use in Rural Communities of the Peruvian Amazon (Iquitos and Vicinity, Maynas Province). Tropical Conservation Science, 382–402. https://doi.org/10.1177/194008291400700303
Terborgh, J. (1988) The big things that run the world — a sequel to E.O. Wilson. Conservation Biology, 2 (4), 402–403.
Terborgh, J., Lopez, L., Nuñez V., P., Rao, M., Shahabuddin, G., Orihuela, G. … Balbas, L. (2002). Ecological Meltdown in Predator-Free Forest Fragments. Science, 294(5548): 1923. Retrieved from https://search.ebscohost.com/login.aspx?direct=true&AuthType=cookie,ip&db=edsjsr&AN=edsjsr.3085421&site=eds-live&scope=site
Turvey, S. T., Fernandez-Secades, C., Nunez-Mino, J. M., Hart, T., Martinez, P., Brocca, J. L., & Young, R. P. (2014). Is local ecological knowledge a useful conservation tool for small mammals in a craribbean multicultural landscape? Biological Conservation, 169, 189-197.
Turvey, S. T., Tien Trung, C., Dai Quyet, V., Van Nhu, H., Van Thoai, D., Cong Anh Tuan, V., Thi Hoa, D., Kacha, K., Sysomphone, T., Wallate, S., Thi Thanh Hai, C., Van Thanh, N., & Wilkinson, N. M. (2015). Interview-based sighting histories can inform regional conservation prioritization for highly threatened cryptic species. Journal of Applied Ecology, 52, 422-433.
Weber, W. & Rabinowitz, A. (1996). A global perspective on large carnivore conservation. Conservation Biology, 10:1046–1054.
Young, A. G. & Clarke, G. M. (2000). Conclusions and future directions: What do we know about genetic and demographic effects of habitat fragmentation and where do we go from here? In: Young, A. G. & Clarke, G. M. (Eds.) Genetics, Demography and Viability of Fragmented Populations. Cambridge University Press, Cambridge, England. Pp. 361-366.
Zeller, K. A., Nijhawan, S., Salom-Perez, R., Potosme, S. H. & Hines, J. E. (2011). Integrating occupancy modeling and interview data for corridor identification: a case for jaguars in Nicaragua. Biological Conservation, 144: 891-901.
9.0 Appendix
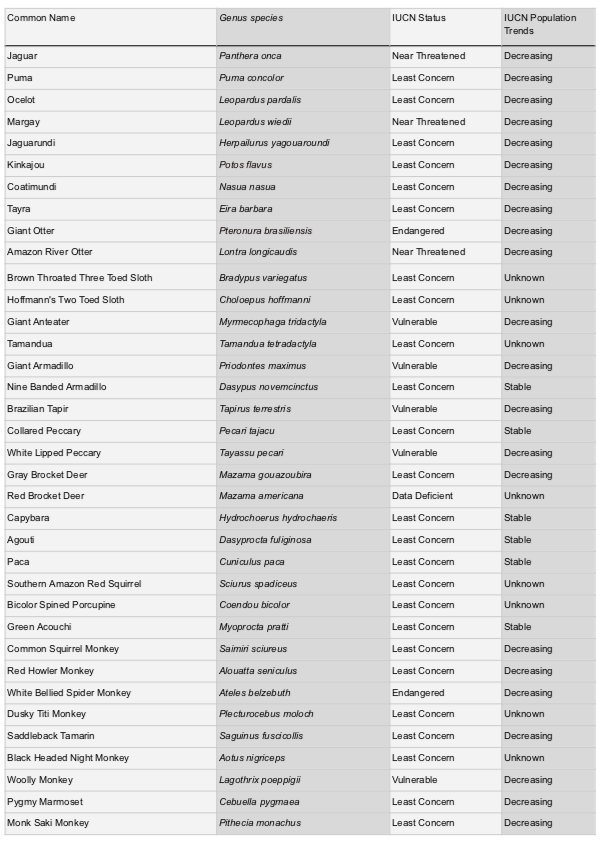
9.1 Basic Information for the 36 Neotropical Mammal Species Surveyed
9.2 Map of the Partnered Adopt-A-School Communities in the Peruvian Amazon Surveyed for Mammals Data
9.3 Communities in Adopt-A-School (Survey Year Data Used and Percent Accuracy from Prior Survey)
03 de Mayo (2019), 11 de Agosto - TBD Ucayali, 28 de Octubre (2019) .916, Auca Cocha (2019), Canada (2018), Canal Pinto (2019), Cedro Isla - TBD Ucayali, Centro Unido (2019) .722, General Merino (2019) .861, Jorge Chavez (2019), Juancho Playa (2019), Juventud Yarina (2019), Las Palmas (2019), Leon Isla (2019), Llachapa (2019), Los Invencibles (2019) .833, Manati I Zona (2019), Miraflores (2019), Nueva Esperanza (2019), Nueva York - TBD Ucayali, Nuevo San Juan del Amazonas (2019), Nuevo Triunfo (2019), Nuevo Triunfo C.A. (2019), Nuevo Uchiza (2019), Nueva Vida (2019), Primero de Enero (2019), Pucallpa (2019), Puerto Huaman (2019), Puerto Rico (2019), Ramon Castilla (2018), San Alejandro (2019), San Antonio de Miraño (2019), San Jose Parananpura - TBD Ucayali, San Juan De Floresta (2018), San Luis (2019), San Pedro de Manati (2019) .916, San Pedro de Mangua (2019), Santa Isabel (2019), Santa Lucia (2019), Santa Maria de Fatima (2019), Santa Victoria (2019), Sucusari (2019) 1.00, Suni Caño (2019), Timicurillo (2019) .722, Timicuro I (2019), Urco Miraño (2019), Yanayacu Timicuro (2019), Yanamono II (2019), Yarina Isla (2019), Yurac Yacu Altura (2019)
9.4 Survey Tool
9.5 Relative Frequency of Mammal Species Detected in 46 Northeast Peruvian Amazon Communities
9.6 Table of Relative Frequency of Mammal Species Presence for Communities Located on the Amazon, Napo, Yanayacu, and Manati River Systems

9.7 Scatter Plot Graph of Relative Frequency of Mammal Species Presence for Communities Located on the Amazon, Napo, Yanayacu, and Manati River Systems

9.8 Reported Commonality of Mammal Species Detection Events (Presence)

9.9 Reported Commonality of Felidae Species Detection Events
9.10 Reported Commonality of Procyonidae Species Detection Events

9.11 Reported Commonality of Mustelidae Detection Events
9.12 Reported Commonality of Xenarthra Species Detection Events
9.13 Reported Commonality of Ungulata Species Detection Events
9.14 Reported Commonality of Rodentia Species Detection Events
9.15 Reported Commonality of Primate Species Detection Events
9.16 Mammal Species Population Trends Over the Past 10 Years as Reported by Community Members

9.17 Correlations Between the Presence of Agricultural Practices and the Quantity of Species Reported Present in Surveyed Communities
